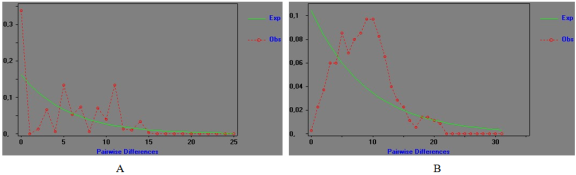
In Senegal, ovarian cancer is the 3rd most common cancer in women with an incidence of 5.0/100,000 women. Thirty-five cancerous tissues, twenty-seven healthy tissues were included in this study. Due to the anatomical position of the ovary, the removal of a sample of suspicious tissue from each patient involves surgery through laparotomy or laparoscopy after obtaining consent. DNA extraction, polymerase chain reaction (PCR) and sequencing were performed to obtain sequences. BioEdit version 7.0.5.3 2005, Harlequin version 3.0, DnaSP version 5.10.01, MEGA 6 were used to perform the analyses. The results show a higher percentage of transition in cancerous tissues (91.45) than in healthy tissues (75.19) in contrast to transversions which are greater in healthy tissues (24.84) than in cancerous tissues (8.54), and the mutation rate (R) is also higher in cancerous tissues (10.712) than in healthy tissues (3.079). Analysis of the polymorphism revealed high values of haplotypic diversity in both cancerous tissues (0.662±0.085) and healthy tissues (0.997±0.011), and low nucleotide diversity values in both tissues (cancerous tissues=0.00922±0.00175; healthy tissues=0.01539±0.00175), these results show us that the genetic evolution of mutations in ovarian cancer has a strong polymorphism. It was also found that the value of the genetic distance between healthy tissues (0.016) was higher than that observed between cancerous tissues (0.009). The genetic distance between healthy and cancerous tissues is 0.015 closer than that observed between healthy tissues. The value of genetic differentiation between healthy and cancerous tissues is significant; this demonstrates a much faster proliferation of cancer cells. The objective of this study is, on the one hand, to better understand the target population by clearly identifying demographic parameters and on the other hand, to evaluate the involvement of somatic mutations and mitochondrial DNA gene expression in the occurrence of ovarian cancer in women in Senegal. The specific objectives are to search for mutations of interest by sequencing mtDNA genes with quasi-maternal inheritance and the impact of these mutations in the D-loop region in healthy and diseased tissues in the patient, but also to learn about the diversity, differentiation and genetic evolution of ovarian cancer in Senegalese women.
| Published in | International Journal of Genetics and Genomics (Volume 12, Issue 4) |
| DOI | 10.11648/j.ijgg.20241204.18 |
| Page(s) | 127-135 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Cancer, Ovary, Mutations, Epidemiology, D-loop, Senegal
Amplified gene | Composants | Volume |
|---|---|---|
D-loop | MilliQ water | 32,9 μl |
Tampon 10X | 5 μl | |
dNTP | 2 μl | |
MgCl2 | 3 μl | |
Primers (sense and nonsense) | 5 μl | |
Polymerase Taq | 0,1 μl | |
DNA extract | 2 μl |
Gene | Primer sequence | Amplification conditions |
|---|---|---|
D-loop | H408 (5’TGTTAAAAGTGCATACCGCCA3’) L16340 (5’AGCCATTTACCGTACATAGCACA3’) | 95°C for 15mn; 35 cycles (95°C for 30s, 62°C for 30s, 72°C for 2mn) 72°C for 10mn |
Variation | Nature | TS | TC | locus | Associated diseases |
|---|---|---|---|---|---|
C33G | Transversion | 1 | ATT | ||
C61T | Transition | 2 | HVS2/ATT | ||
C64T | Transition | 2 | HVS2/ATT | ||
A73G | Transition | 25 | 35 | HVS2/ATT | |
T89C | Transition | 20 | HVS2/ATT | ||
A93G | Transition | 3 | 24 | HVS2/ATT | |
A95C | Transversion | 2 | 2 | HVS2/ATT | |
G97A | Transition | 20 | HVS2/ATT | ||
C114G | Transversion | 1 | HVS2/OH/ATT | ||
G143A | Transition | 3 | HVS2/OH/ATT | ||
T146C | Transition | 6 | 26 | HVS2/OH/ATT | Absence of endometriosis |
C150T | Transition | 10 | 30 | HVS2/OH/ATT | Conflicting reports longevity/cervical carcinoma/risk of HPV infection |
C151T | Transition | 3 | 1 | HVS2/OH/ATT | |
C151G | Transversion | 1 | HVS2/OH/ATT | ||
T152C | Transition | 15 | 33 | HVS2/OH/ATT | |
C182T | Transition | 7 | 31 | HVS2/OH/ATT | |
G185T | Transversion | 3 | 2 | HVS2/OH/ATT | |
G185C | Transversion | 2 | HVS2/OH/ATT | ||
C186A | Transversion | 1 | 1 | HVS2/OH/ATT | |
A189C | Transversion | 1 | 1 | HVS2/OH/ATT | |
A189G | Transition | 1 | HVS2/OH/ATT | ||
T195C | Transition | 14 | 32 | HVS2/OH/ATT | Melanoma/MB-associated points |
A197C | Transversion | 1 | HVS2/OH/ATT | ||
C198T | Transition | 6 | 30 | HVS2/OH/ATT | |
T204C | Transition | 4 | 6 | HVS2/OH/ATT | |
T204G | Transversion | 1 | HVS2/OH/ATT | ||
G207A | Transition | 4 | 2 | HVS2/OH/ATT | |
G221C | Transversion | 1 | HVS2/OH/ATT | ||
Variation | Nature | TS | TC | Locus | Associated diseases |
A232C | Transversion | 1 | HVS2/OH/ATT | ||
A240T | Transversion | 1 | HVS2/OH/ATT | ||
G247A | Transition | 3 | 3 | HVS2/OH/ATT | |
C256T | Transition | 1 | HVS2/OH/ATT | ||
A263G | Transition | 27 | 35 | HVS2/OH/ATT | |
T279C | Transition | 1 | HVS2/OH/ATT | ||
T292A | Transversion | 1 | HVS2/OH/ATT | ||
A297G | Transition | 1 | HVS2/OH/ATT | ||
C309T | Transition | 1 | HVS2/OH/ATT | ||
C315CC | Insertion | 26 | 34 | HVS2/OH/ATT | Melanoma patients |
G316C | Transversion | 1 | 1 | HVS2/OH/ATT | |
G316GA | insertion | 1 | 1 | HVS2/OH/ATT | |
C325T | Transition | 26 | HVS2/OH/ATT | ||
C332T | Transition | 1 | HVS2/OH/ATT | ||
A357G | Transition | 3 | 2 | HVS2/OH/ATT | |
T16386A | Transversion | 2 | ATT | ||
G16390A | Transition | 7 | 28 | ATT | POAG-association potential |
C16395G | Transversion | 1 | ATT | ||
C16411A | Transversion | 1 | ATT | ||
A16421C | Transversion | 1 | ATT | ||
T16422C | Transition | 1 | ATT | ||
T16468G | Transversion | 1 | ATT | ||
T16469G | Transversion | 2 | ATT | ||
G16477A | Transition | 1 | ATT | ||
C16478A | Transversion | 3 | ATT | ||
T16479A | Transversion | 1 | ATT | ||
C16501G | Transversion | 1 | ATT | ||
C16511T | Transition | 1 | ATT | ||
T16519C | Transition | 19 | 27 | ATT | |
T16522A | Transversion | 1 | ATT | ||
A16525G | Transition | 1 | ATT | ||
C16527T | Transition | 2 | ATT |
Parameters | Cancerous tissues | Healthy tissues |
|---|---|---|
Number of sequences | 35 | 27 |
Number of haplotypic | 8 | 26 |
Number of sites | 559 | 559 |
Number of monomorphic sites | 536 | 509 |
Number of polymorphic sites | 23 | 50 |
Number of sparing information sites | 19 | 23 |
Nucleotidic frequence | A G T C 24.91 30.20 27.51 17.38 | A G T C 24.87 30.20 27.46 17.47 |
% Transitions | 91.45 | 75.19 |
% Transversions | 8.54 | 24.84 |
Rate of Transitions/Transversions=R | 10.712 | 3.079 |
Haplotypic diversity (HD±SD) | 0.662±0.085 | 0.997±0.011 |
Nucleotide diversity (Pi±SD) | 0.00922±0.00175 | 0.01539±0.00175 |
average number of nucleotide differences (K) | 42.581 | 11.901 |
Tissues | Genetic distance intra | Genetic distance inter | Fst (P-value) |
|---|---|---|---|
Cancerous | 0.009±0.002 | 0.015±0.003 | 0.333 (P=0.000) |
Healthy | 0.016±0.003 |
Parameters | Healthy Tissues | Cancerous Tissues |
|---|---|---|
D of Tajima | -1,278 | -0,266 |
P-value | 0,081 | 0,443 |
Fs of Fu | -19,113 | 2,590 |
DNA | DeoxyriboNucleic Acid |
WHO | World Health Organization |
FIGO | International Federation of Gynecology and Obstetrics |
HAH | High Authority of Health |
mtDNA | Mitochondria DeoxyriboNucleic Acid |
FST | Degree of Genetic Differentiation or Fixation Index |
D-LOOP | Displacement Loop |
PCR | Polymerase Chain Reaction |
DNAsp | DeoxyriboNucleic Acid Sequence Polymorphism |
MEGA | Molecular Evolutionary Genetics Analysis |
AMOVA | Analysis of Molecular Variance |
SSD | Sum of Squares Deviation |
RI | Raggedness Index |
HVS | Hypervariable Segments |
| [1] | Synthese_rapport_activite_2010 |
| [2] | Negrini S, Gorgoulis VG, Halazonetis TD. Instabilité génomique – une caractéristique évolutive du cancer. Nat Rev Mol Cell Biol. 2010; 11: 220–228. |
| [3] | Vasileiou, PVS; Mourouzis, I.; Pantos, C. Principaux aspects concernant le maintien de l'intégrité du génome mitochondrial des mammifères. Int. J. Mol. Sci. 2017, 18, 1821. |
| [4] | Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. InNucleic acids symposium series 1999 Jan 1 (Vol. 41, No. 41, pp. 95-98). [London]: Information Retrieval Ltd., c1979 c2000. |
| [5] | Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research. 1994 Nov 11; 22(22): 4673-80. |
| [6] | RM Andrews, Je Kubacka, PF Chinnery, Les hiboux-lumière de la RN, DM Turnbull, N. Howell: Réanalyse et révision de la séquence de référence de Cambridge pour l'ADN mitochondrial humain. 1999 octobre; 23(2): 147. |
| [7] | Rozas, J., Librado, P., Sánchez-Del Barrio, J. C., Messeguer, X. and Rozas, R. (2012) DnaSP Version 5 Help Contents [Help File]. |
| [8] | Tamura K, Stecher G, Peterson D, et al. MEGA6 Analyse génétique évolutive moléculaire Version 6.0. Mol Biol Evol. 2013. 30: 2725-2729. |
| [9] | Nei M. & Gojobori T., (1986): Simple methods for estimating the numbers of synonymous & nonsynonymous nucleotide substitutions. Molecular Biology & Evolution (3): 418-426. |
| [10] | B. Weir, WG Hill. Estimation des statistiques F. Publié dans la Revue annuelle de génétique 2002. |
| [11] | Excoffier L, Estoup A, Cornuet J-M (2005) Bayesian Analysis of an Admixture Model With Mutations and Arbitrarily Linked Markers. Genetics 169: 1727-1738. |
| [12] | Tajima, F. 1989a. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 123:585-595, 597-601. |
| [13] | Fu, Y.-X. (1997) Statistical tests of neutrality of mutations against population growth, hitchhiking and backgroud selection. Genetics 147: 915-925. |
| [14] | Guerra F, Kurelac I, Magini P, et al. Mitochondrial DNA genotyping reveals synchronous nature of simultaneously detected endometrial and ovarian cancers. Gynecol Oncol. 2011; 122(2): 457-458. |
| [15] | Calloway CD, Reynolds RL, Herrin Jr GL, Anderson WW. The frequency of heteroplasmy in the HVII region of mtDNA differs across tissue types and increases with age. The American Journal of Genetics. 2000 Apr 1; 66(4): 1384-1397. |
| [16] | Li H, Hong ZH. Mitochondrial DNA mutations in human tumor cells. Oncology Letters. 2012 Nov 1; 4(5): 868-872. |
| [17] | Brandon M, Baldi PA, Wallace DC. Mitochondrial mutations in cancer. Oncogene. 2006 Aug; 25(34): 4647- 62. |
| [18] | Montserrat Gómez-Zaera, Jésus Avril, Laura Gonzalez, Fernando Aguilo, Préservatif Enric, Marga Nadal, Virgine Nunes, Recherche sur les mutations/Mécanismes fondamentaux et moléculaires de la mutagenèse Volume 595, numéros 1–2, 20 mars 2006, pages 42 51. |
| [19] | Tan DJ, Bai RK, Wong LJ. Comprehensive scanning of somatic mitochondrial DNA mutations in breast cancer. Cancer Res. 2002; 62(4): 972-976. |
| [20] | Tan et al. Nouveaux décalages hétéroplasmiques du cadre de lecture et mutations faux-sens de l'ADN mitochondrial somatique dans le cancer de la bouche des mâcheurs de chique de bétel Gènes Chromosomes Cancer (2003). |
| [21] | Bragoszewski P, Kupryjanczyk J, Bartnik E, Rachinger A, Ostrowski J. Limited clinical relevance of mitochondrial DNA mutation and gene expression analyses in ovarian cancer. BMC Cancer. 2008 Dec 1; 8(1): 292. |
| [22] | Chen JZ, Gokden N, Greene GF, Mukunyadzi P, Kadlubar FF. Extensive somatic mitochondrial mutations in primary prostate cancer using laser capture microdissection. Cancer Research. 2002 Nov 15; 62(22): 6470-6474. |
| [23] | Doupa D. Benign Breast Tumors among Senegalese Women: Diversity and Genetic Evolution of D-Loop. Open Access Library Journal. 2015; 2(08): 1. |
| [24] | M. Kimura. The Neutral Theory of Molecular Evolution. Cambridge University Press, Cambridge, UK, 1983. |
| [25] | Nowell PC. The clonal evolution of tumor cell populations. Science 1976; 194: 23–28. |
| [26] | M. Kimura. Evolutionary rate at the molecular level. Nature, 217: 624–26, 1968. |
APA Style
Fall, H., Mbaye, F., Sembene, M. (2024). Impact of Mutations in the D-loop Region in Ovarian Cancer in Senegalese Women. International Journal of Genetics and Genomics, 12(4), 127-135. https://doi.org/10.11648/j.ijgg.20241204.18
ACS Style
Fall, H.; Mbaye, F.; Sembene, M. Impact of Mutations in the D-loop Region in Ovarian Cancer in Senegalese Women. Int. J. Genet. Genomics 2024, 12(4), 127-135. doi: 10.11648/j.ijgg.20241204.18
AMA Style
Fall H, Mbaye F, Sembene M. Impact of Mutations in the D-loop Region in Ovarian Cancer in Senegalese Women. Int J Genet Genomics. 2024;12(4):127-135. doi: 10.11648/j.ijgg.20241204.18
@article{10.11648/j.ijgg.20241204.18,
author = {Habib Fall and Fatimata Mbaye and Mbacké Sembene},
title = {Impact of Mutations in the D-loop Region in Ovarian Cancer in Senegalese Women
},
journal = {International Journal of Genetics and Genomics},
volume = {12},
number = {4},
pages = {127-135},
doi = {10.11648/j.ijgg.20241204.18},
url = {https://doi.org/10.11648/j.ijgg.20241204.18},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijgg.20241204.18},
abstract = {In Senegal, ovarian cancer is the 3rd most common cancer in women with an incidence of 5.0/100,000 women. Thirty-five cancerous tissues, twenty-seven healthy tissues were included in this study. Due to the anatomical position of the ovary, the removal of a sample of suspicious tissue from each patient involves surgery through laparotomy or laparoscopy after obtaining consent. DNA extraction, polymerase chain reaction (PCR) and sequencing were performed to obtain sequences. BioEdit version 7.0.5.3 2005, Harlequin version 3.0, DnaSP version 5.10.01, MEGA 6 were used to perform the analyses. The results show a higher percentage of transition in cancerous tissues (91.45) than in healthy tissues (75.19) in contrast to transversions which are greater in healthy tissues (24.84) than in cancerous tissues (8.54), and the mutation rate (R) is also higher in cancerous tissues (10.712) than in healthy tissues (3.079). Analysis of the polymorphism revealed high values of haplotypic diversity in both cancerous tissues (0.662±0.085) and healthy tissues (0.997±0.011), and low nucleotide diversity values in both tissues (cancerous tissues=0.00922±0.00175; healthy tissues=0.01539±0.00175), these results show us that the genetic evolution of mutations in ovarian cancer has a strong polymorphism. It was also found that the value of the genetic distance between healthy tissues (0.016) was higher than that observed between cancerous tissues (0.009). The genetic distance between healthy and cancerous tissues is 0.015 closer than that observed between healthy tissues. The value of genetic differentiation between healthy and cancerous tissues is significant; this demonstrates a much faster proliferation of cancer cells. The objective of this study is, on the one hand, to better understand the target population by clearly identifying demographic parameters and on the other hand, to evaluate the involvement of somatic mutations and mitochondrial DNA gene expression in the occurrence of ovarian cancer in women in Senegal. The specific objectives are to search for mutations of interest by sequencing mtDNA genes with quasi-maternal inheritance and the impact of these mutations in the D-loop region in healthy and diseased tissues in the patient, but also to learn about the diversity, differentiation and genetic evolution of ovarian cancer in Senegalese women.
},
year = {2024}
}
TY - JOUR T1 - Impact of Mutations in the D-loop Region in Ovarian Cancer in Senegalese Women AU - Habib Fall AU - Fatimata Mbaye AU - Mbacké Sembene Y1 - 2024/11/29 PY - 2024 N1 - https://doi.org/10.11648/j.ijgg.20241204.18 DO - 10.11648/j.ijgg.20241204.18 T2 - International Journal of Genetics and Genomics JF - International Journal of Genetics and Genomics JO - International Journal of Genetics and Genomics SP - 127 EP - 135 PB - Science Publishing Group SN - 2376-7359 UR - https://doi.org/10.11648/j.ijgg.20241204.18 AB - In Senegal, ovarian cancer is the 3rd most common cancer in women with an incidence of 5.0/100,000 women. Thirty-five cancerous tissues, twenty-seven healthy tissues were included in this study. Due to the anatomical position of the ovary, the removal of a sample of suspicious tissue from each patient involves surgery through laparotomy or laparoscopy after obtaining consent. DNA extraction, polymerase chain reaction (PCR) and sequencing were performed to obtain sequences. BioEdit version 7.0.5.3 2005, Harlequin version 3.0, DnaSP version 5.10.01, MEGA 6 were used to perform the analyses. The results show a higher percentage of transition in cancerous tissues (91.45) than in healthy tissues (75.19) in contrast to transversions which are greater in healthy tissues (24.84) than in cancerous tissues (8.54), and the mutation rate (R) is also higher in cancerous tissues (10.712) than in healthy tissues (3.079). Analysis of the polymorphism revealed high values of haplotypic diversity in both cancerous tissues (0.662±0.085) and healthy tissues (0.997±0.011), and low nucleotide diversity values in both tissues (cancerous tissues=0.00922±0.00175; healthy tissues=0.01539±0.00175), these results show us that the genetic evolution of mutations in ovarian cancer has a strong polymorphism. It was also found that the value of the genetic distance between healthy tissues (0.016) was higher than that observed between cancerous tissues (0.009). The genetic distance between healthy and cancerous tissues is 0.015 closer than that observed between healthy tissues. The value of genetic differentiation between healthy and cancerous tissues is significant; this demonstrates a much faster proliferation of cancer cells. The objective of this study is, on the one hand, to better understand the target population by clearly identifying demographic parameters and on the other hand, to evaluate the involvement of somatic mutations and mitochondrial DNA gene expression in the occurrence of ovarian cancer in women in Senegal. The specific objectives are to search for mutations of interest by sequencing mtDNA genes with quasi-maternal inheritance and the impact of these mutations in the D-loop region in healthy and diseased tissues in the patient, but also to learn about the diversity, differentiation and genetic evolution of ovarian cancer in Senegalese women. VL - 12 IS - 4 ER -
Genomics Laboratory, Department of Animal Biology, Faculty of Science and Technology, Cheikh Anta Diop University, Dakar, Senegal
Genomics Laboratory, Department of Animal Biology, Faculty of Science and Technology, Cheikh Anta Diop University, Dakar, Senegal
Genomics Laboratory, Department of Animal Biology, Faculty of Science and Technology, Cheikh Anta Diop University, Dakar, Senegal